Corneliu Neamțu este profesor de fiziologie la UMF „Gr. T.
Popa” Iași și este prim-vicepreședinte al Asociației de Algeziologie din
România.
Durerea reprezintă
motivul cel mai frecvent pentru care bolnavul solicită ajutor medicului,
aproape neexistând specialitate medicală care să nu se confrunte cu prezența
unei simptomatologii dureroase. Între anumite limite ea reprezintă un fenomennormal de semnalare a unui pericol din mediul extern, dar și intern al
omului, privind debutul unui proces ostil în organism care tinde să îi tulbure
homeostazia. În această interpretare, prezența durerii este fiziologică,
utilă și benefică, constituind un factor esențial de asigurare a supraviețuirii
și se referă la tot ce înțelegem terminologic prin durerea acută.
Disocierea între
aspectul util al durerii și cel generator al stării de boală este esențială
pentru practica medicală. „Durerea alarmează bolnavul, dar liniștește pe
medic”, constituind un semn etiologic, topografic sau chiar patogenic, cu
importanță relevantă uneori în stabilirea unui diagnostic curent și a unei
strategii terapeutice adecvate. În schimb, durerea cronică, chinuitoare,
progresivă, are zi de zi efecte destructive și chiar epuizante (fizice,
funcționale, endocrine, metabolice, neuropsihice), care uneori pot fi cu greu
depășite și pot evolua până la invalidarea pacientului.
Fenomenului dureros i se
pot distinge patru componente principale: senzitivo-senzorială discriminativă
(localizare, natura durerii, intensitate, durată); psihică
motivațional-emoțională (descrierea extrem de variabilă simbolic-metaforică a
durerii, cu un spectru semantic larg și diferit de la individ la individ
asociat cu reacții gestuale); componenta psihică cognitivă și evaluativă, ce
implică fenomene de atenție, analiză și comparație cu alte experiențe similare
anterioare și de anticipare a riscurilor (1, 2); componenta motorie
somato-vegetativă ca model de răspuns reflex asociat (reacția la durere).
Durerea declanșează o
adevărată cascadă de procese fiziologice, de la recepție până la interpretare
și reacție, etapele fiind codificate, având în final o rezultantă integrată a
tuturor interfețelor complementare și cooperante. Procesele fiziologice
declanșate, cu răsunet neuroendocrin, metabolic și chiar genetic (ultimele
prezente chiar și în durerea de durată scurtă cum sunt creșterea
protooncogenelor c-fos neuronale și modificări ale transcripției în
neuronii nociceptivi, de exemplu) (3) modifică profund homeostazia
organismului și pot fi evaluate prin măsurători clinice și instrumentale.
Manifestările observate
în durere pot cunoaște variabile cantitativ-calitative, clasificate de modelul
Fordyce (4) în patru clase fundamentale: nocicepția (răspunsurile
sistemului nervos la impulsuri neplăcute, aversive); durerea (percepția
conștientizată a aferențelor dureroase); suferința (răspunsurile
emoționale negative la nocicepție și durere); comportamentul la durere
(comunicări motorii, verbale, gestuale, expresionale). Comportamentul la
durere, ultimul de pe această scală graduală, evidențiază manifestări complexe
care pot fi controlate prin reacții reflexe posturale (antalgice) sau motorii
(masajul, mersul), umorale (intervenția opioidelor endogene, a serotoninei și a
dopaminei) sau psihologic (autocontrolul bazat pe cunoaștere și exerciții de
reglare și armonizare a propriului comportament). Sistemele de control se
realizează prin bucle anticipative (feed-forward) sau prin bucle de
reacție inversă (feedback). Activarea unui control psihologic al
comportamentului la durere a restructurat în ultimul timp managementul suferinței
dureroase în clinicile de terapie a durerii, încurajând atitudinile pozitive și
ajutând pacientul să învețe tehnici de automonitorizare, de relaxare, crescând
capacitatea acestuia de a-și controla durerea (5).
În
prezent, conceptul ce privește durerea ca fenomen multidimensional are la bază
existența dovedită a unor rețele de structuri neuroanatomice și neurochimice de
lucru (inclusiv endocrine), cu rol fundamental în convertirea senzațiilor
nociceptive în percepții și trăiri ale durerii, care se deosebesc profund din
acest punct de vedere de alte sensibilități. Ansamblul de rețele ale durerii
prezente în sistemul nervos este cunoscută sub numele de neuromatricea durerii
(6). Principalele structuri implicate sunt: ariile spinale, talamus, nucleul lentiform,
girusul cingulat, girusul prefrontal, lobul insular, cortexul inferior parietal
și somato-senzorial (7).
Ca model cerebral
analitic, durerea este rezultatul unei suprasumații de stimuli (algogeni) de
naturi variate, vehiculați și mediați ascendent prin sisteme de centri și căi
complexe specifice și nespecifice, având ca țintă de proiecție structuri
cerebrale psiho-somato-vegetative. Pe parcursul acestor sisteme au loc recepția
și transmisia semnalelor nociceptive, procesarea acestora (analiza, codificarea,
integrarea) și modularea durerii.
Legătura cauzală între
leziunea organică și durere este frecventă, dar nu obligatorie, putând exista
leziune fără durere și durere fără leziune (durerile psihoafective). Totuși,
într-o măsură copleșitoare, prezența durerii este un semn important al
existenței unei leziuni organice sau funcționale.
Stimulii algogeni
Din punct de vedere
sistematic, clasificarea agenților algogeni cuprinde stimuli fizici, chimici și
biologici. Deși stimulii fizici, frecvent întâlniți în durerea fiziologică și
reprezentați de stimuli mecanici (atingere, presiune, vibrație, tracțiune,
distensie, înțepare), excită simultan, atât receptorii pentru durere
(nociceptorii) cu prag înalt, cât și mecanoreceptori (receptorii tactili) cu
prag scăzut, descărcările mecano-receptoare se adaptează mai rapid decât
nociceptorii, iar raportul aferențelor nociceptori/receptori tactili se schimbă
în favoarea nociceptorilor. În durerea patologică, modelul de stimulare
mecanică este reprezentat de distensia organelor cavitare și a capsulei
organelor parenchimatoase, de tracțiunea ligamentelor, distensiile vasculare,
vasoconstricție (spasm, ischemie) sau vasodilatație asociate cu compresiunea
terminațiilor nervoase.
Pragul mecanic pentru
stimularea nociceptorilor este de 6 ± 0,6 bari pentru receptorii
terminațiilor fibrelor C și 3,5 ± 0,3 bari pentru fibrele A-delta.
Stimulii termici
aplicați în mod obișnuit asupra pielii pot acționa prin temperaturi scăzute
care activează nociceptori profunzi din marile vase venoase. Durerea este
intensă și este urmarea vasoconstricției cutanate. Durerea este stimulată și de
energia radiantă, hipertermică, când temperatura aplicată atinge 45°C.
Stimulii chimici sunt
frecvent întâlniți în durerea patologică. De exemplu, durerea postischemică
apare ca efect al acidozei prin hipoxie, ea fiind percepută de la un pH de 6,2
cu un maximum de intensitate la pH de 3,2. Dintre agenții chimici mai frecvent
algostimulatori se notează: aminele (serotonina, histamina, acetilcolina), monoxidul
de azot, peptide (bradikinina, substanța P), prostaglandine și ionii de H+,
K+.
Pragul la durere
Pragul la durere este un
test subiectiv reprezentat de prima senzație dureroasă provocată de un stimul
cu intensitate minimă și pe care subiectul uman o poate semnaliza verbal.
Măsurarea pragului se face prin tatonare și evaluarea intensității celei mai
reduse a stimulului algogen care evocă pragul. Dificultățile în evaluarea
pragului la durere se datorează în primul rând caracterului multidimensional al
percepției dureroase (senzorial-discriminativă, afectiv-emoțională și
cognitiv-discriminativă), ceea ce face din interpretarea și trăirea durerii un
proces particularizat și chiar personalizat de la individ la individ.
Receptorii pentru durere
(nociceptorii) se clasifică în receptori monomodali (specifici) activați
de stimuli mecanici și polimodali (nespecifici) activați de stimuli
mecanici, termici, chimici și biologici. Pragul de activare al nociceptorilor
polimodali este diferit de cel al receptorilor monomodali, fiind dependent de
calitatea stimulului mecanic, iar excitațiile nenociceptive ale
mecanoreceptorilor și termoreceptorilor sunt recepționate la un nivel mai
coborât al intensității stimulilor decât cele nociceptive (8). Creșterea
intensității stimulului algogen induce în fibrele nociconducătoare A-delta și C
potențiale de acțiune a căror frecvență crește proporțional cu intensitatea
stimulului aplicat. Limitele pragului la durere sunt înguste, durerea apărând
la o intensitate a stimulului egală cu de două-trei ori mai mare decât
intensitatea stimulului senzorial. Receptorii polimodali ai fibrelor
algoconducătoare C pot fi hipersensibilizați de stimulări repetitive dureroase,
fiind capabili să realizeze sumarea (convergența) spațială atât la periferie,
cât și în ariile neuronale centrale. Nociceptorii monomodali au un prag de
excitabilitate la durere mai înalt, în timp ce nociceptorii polimodali au un
prag mai scăzut. Pragul la care nociceptorii încep să descarce biopotențiale de
acțiune nu coincide în mod obișnuit cu pragul percepției durerii la om și cu
pragul reacției la durere la animale. De cele mai multe ori, pragul la durere
este mai înalt decât pragul de nociceptor.
Cel mai potrivit răspuns
al organismului în fața durerii este cel care oferă adaptarea optimă la
eventuale condiții ostile ale mediului extern sau intern și acesta este cazul
îndeosebi al durerii acute. Durata prelungită în timp a durerii și
transformarea ei din semnal și simptom în sindrom sau boală, cum este cazul
durerii cronice, evidențiază situația în care sistemele proprii reactive, de
atenuare și adaptare, sunt depășite și ele trebuie corectate și recuperate prin
agenți terapeutici fizici, chimici, aloterapie și/sau psihoterapie.
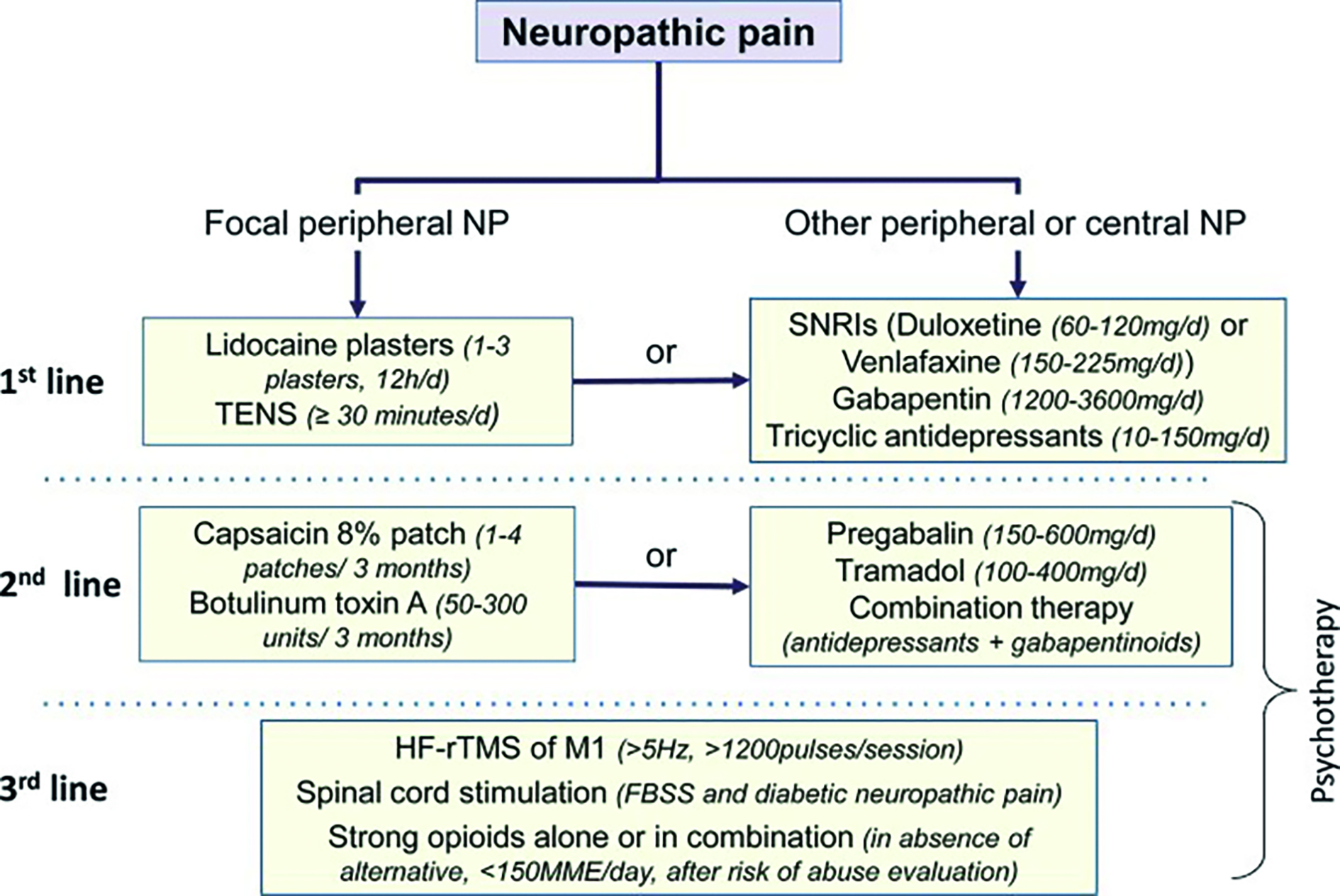
Tipuri de durere
Durerea este clasificată
în două tipuri distincte: rapidă (primară) și întârziată/lentă (secundară).
Durerea rapidă are loc la aproximativ 0,1 sec din momentul aplicării stimulului
dureros, în timp ce durerea lentă apare la interval de o secundă sau mai mult,
crescând lent timp de câteva secunde sau chiar minute. Durerea rapidă are ca
prototip senzația de înțepătură, este percepută ca ascuțită, este bine
localizată, resimțită rapid și fugace, durează atâta timp cât acționează
stimulul, fiind dependentă de fibrele nervoase A-delta. Durerea lentă include
prezența de leziuni tisulare, cu afectarea integrității anatomofuncționale a
receptorilor și a căilor de conducere. Apare ca urmare a unor procese
biochimice locale care eliberează substanțe endogene cu un puternic efect
dureros: ioni de K+, prostaglandina E, bradikinină, substanța P,
histamina, serotonină, monoxidul de azot. Acest tip de durere este imprecis
descris ca localizare, are conducere lentă, durează și după încetarea acțiunii
stimulului algogen, declanșează reacții musculare tonice.
Recepția durerii
Clasificarea
nociceptorilor pe criterii funcționale îi împarte în: nociceptori
mecano-senzitivi atașați fibrelor algo-conductoare A-delta; nociceptori
mecano-termici atașați fibrelor A-delta; nociceptori polimodali, care răspund
la stimuli termici, mecanici și chimici, aparținând fibrelor algoconductoare C.
Primele două categorii corespund durerii rapide, iar a treia exprimă durerea
difuză, profundă, lentă.
Din punct de vedere
morfologic, toți receptorii durerii sunt terminații nervoase libere, dar nu
este exclusă și recepția durerii de terminații capsulate, necapsulate și rețele
terminale perivasculare sau din diverse alte țesuturi (periost, suprafețe
articulare). Terminațiile nervoase libere prezente în derm și epiderm sunt în
majoritate mielinizate. Cele neîncapsulate (Merkel și Ruffini) sunt corelate în
special cu schimbările de poziție în țesuturi, iar cele încapsulate (Meissner,
Krause și Vater-Pacini) cu senzațiile de tact și presiune. Se consideră că
receptorii periferici adecvat stimulați induc răspunsuri adecvate de
temperatură, presiune și durere, ultima senzație fiind corelată specific cu
terminațiile nervoase libere. Cu toate acestea, nu se poate afirma încă fără
echivoc că numai terminațiile nervoase libere sunt receptori specifici ai
durerii, deși dacă s-ar respinge teoria specificității ar fi greu de înțeles
rezultatele terapeutice obținute prin înlăturarea neurochirurgicală a tractelor
algoconductoare. Importantă este și relația constantă și direct proporțională
dintre gradul de activare al unui receptor presupus algosensibil și gradul de
intensitate al durerii resimțite, astfel că orice receptor și orice fibră
nervoasă senzitivo-conductoare este capabilă să transmită senzații dureroase,
fiecare clasă de receptori senzitivi și fiecare fibră având nivelul de
intensitate a stimulului aplicat de la care începe transmisia algogenă. În
aceste condiții, dacă există un factor specific, el este reprezentat de
intensitatea adecvată („specifică”) tipului de receptor la care apare durerea
la un individ aflat într-o anumită stare psihoemoțional-afectivă,
sănătos/bolnav, reactiv/areactiv etc.
Stimulii nociceptivi
activează numeroși neuroni și nuclei pe tot parcursul traseului căilor
algoconductoare spinocorticale ascendente, de transport și proiecție a
informațiilor nociceptive, la care se asociază structuri nervoase responsabile
de memoria de lungă durată care servește de comparator al experienței
emoționale dureroase prezente. Acest fapt contribuie decisiv la caracterul multidimensional
al durerii și la procesarea senzațiilor algice în neuromatricea durerii.
Căile de conducere și
releele durerii. Procesarea durerii
Informațiile senzitive
dureroase sunt conduse sub formă de descărcări ale potențialelor de acțiune
nociceptive, de la receptorii periferici până la ariile de proiecție corticale,
pe căile extranevraxiale și intranevraxiale.
Căile aferente
extranevraxiale ale durerii somatice (superficiale și profunde) și viscerale. Fibrele algoconductoare ale
durerii superficiale cutanate sunt: fibre mielinice subțiri cu diametrul de 2–5
μm (grupa fibrelor A-delta și gamma), cu viteza de conducere de 4–30 m/sec
pentru durerea tolerabilă, relativ bine localizată (tip „înțepătură”); fibre
amielinice subțiri, cu diametrul de 0,4–1,2 μm (grupa C), cu viteza de 0,4
m/sec, pentru durerea intensă și difuză (tip „arsură”). Fibrele A-delta și C nu
sunt exclusiv algoconductoare, ele putând conduce și sensibilitățile tactilă,
termică și presională. Fibrele care mediază durerea superficială/cutanată de la
nivelul trunchiului și membrelor intră în structura nervilor spinali fiind
distribuite tegumentar pe arii delimitate topografic numite dermatoame, iar
pentru extremitatea cefalică distribuția aparține teritoriilor nervilor
cranieni senzitivi.
Căile intranevraxiale
ale durerii somatice (superficiale și profunde) și viscerale. Căile senzațiilor dureroase
și termice recepționate din trunchi, membre și din aria cefalică, după
pătrunderea în măduva spinării se înmănunchiază sub forma căii comune anterolaterale
(CCAL) dispusă în cuadrantul cu același nume al substanței albe din măduva
spinării. CCAL este format din trei tracturi ascendente conducătoare ale
influxurilor nociceptive: spino-talamic lateral (tractul neo-spinotalamic,
sistemul lateral al durerii); spino-mezencefalic; spino-reticular (tractul
paleo-spino-talamic, sistemul paramedian, sistemul medial). La acestea se mai
adaugă o cale asociată spino-cervico-talamică, încorporată în coloana dorsală a
sistemului lemniscal al sensibilităților generale (presiune, tact, vibrații,
stereognozia, kinestezia).
Tractul spino-talamic
lateral (TSTL). Fibrele A-delta și C intră ca axoni ai ganglionilor spinali
în măduva spinării prin partea laterală a rădăcinii dorsale a nervilor
rahidienii (schema). Ele se bifurcă într-un ram ascendent și unul
descendent și apoi pătrund și se finalizează sinaptic în cornul posterior
medular, în neuronii laminelor Rexed I, II, V, VI și VII. Fibrele A-delta
realizează sinapse excitatorii cu neuronii senzitivi medulari de proiecție din
lamina I, II și V Rexed, iar fibrele C urmează două variante: sinapsă cu
neuronii senzitivi I, II Rexed din cornul posterior, ai căror axoni vor avea un
traiect ascendent spre centrii nervoși superiori, respectiv sinapsă cu
interneuronii inhibitori care modulează influxurile nociceptive în drumul lor
spre centrii nervoși superiori.
Cea mai importantă din
punctul de vedere al aferențelor dureroase este lamina I, în care se află trei
tipuri de neuroni: nociceptivi specifici (monomodali), care primesc informații
nociceptive cutanate (mecanice sau termice), musculare, articulare și
viscerale; specifici (monomodali), care răspund la stimuli termici nedureroși
pentru frig; nociceptivi polimodali, care transmit în cea mai mare parte prin
fibre C algoconductoare informații dureroase termice (frig, cald), mecanice tip
„ciupitură” și răspund la stimuli musculari, articulari sau viscerali.
În lamina IV și V, loc
de origine a tractului spino-talamic anterior (cale tactilă), există și neuroni
nociceptivi care sunt sensibili la o gamă largă de stimuli nociceptivi și
nenociceptivi aplicați pe arii cutanate extinse și în manieră gradată.
Fibrele A-delta și C
realizează sinapse în măduva spinării la niveluri succesive. În neuronii
cornului posterior (neuroni de ordinul I) are loc procesarea informațiilor
nociceptive și tot aici fac sinapsă fibrele sistemelor descendente modulatoare
ale durerii cu origine în structuri cu rol inhibitor/excitator existente la
diferite etaje ale sistemului nervos central supraspinal. Din neuronii cornului
medular posterior emerg axoni (din care 90% se încrucișează) ce vor conduce
ascendent prin tractul spino-talamic lateral (neo-spino-talamic) influxurile
nociceptive la etajul diencefalic (talamo-hipotalamic) unde se află neuronul III
de pe căile algoconductoare. Fibrele conductoare ale senzațiilor dureroase și
termice ce aduc informații de la corp și membre sunt proiectate în neuroni
talamici din nucleul ventro-postero-lateral, care împreună cu nucleul
ventro-postero-medial ce primește aferențe nociceptive cefalice formează
complexul ventro-bazal, nucleul posterior și nucleii intralaminari. Complexul
ventro-bazal și nucleul posterior primesc descărcări de impulsuri nociceptive
de la neuronii laminelor spinale I, II și V pe cale neo-spino-talamică
(sistemul lateral al durerii), iar la nucleii intralaminari sosesc informații
de la neuronii nociceptivi ai laminelor spinale VI, VII și VIII pe calea
tractului spino-reticulo-talamic (paleo-spino-talamic). Tracturile
spino-talamice sunt organizate somatotopic ascendent pe măsură ce primesc
aferențe de la segmentele corporale, diferențiat, din arii determinate
topografic: dermatoame (somatice) și sclerotoame (viscerale). Plecați din
neuronii nucleului talamic ventro-postero-lateral, care este organizat de
asemenea somatotopic, axonii neuronilor termo-nociceptivi de ordinul III
străbat capsula internă și corona radiata, finalizându-se în lobul parietal al
cortexului cerebral, în lamina IV aria somato-senzitivă a girusului postcentral
(câmpurile 3, 1, 2 din clasificarea Brodmann). Axonii cu origine în nucleul
talamic posterior ajung în lamina IV a ariei somato-senzitive II (câmpurile 3,
1, 2) lângă scizura lui Sylvius.
Zonele din ariile
senzitive corticale sunt esențiale pentru capacitatea de diferențiere și
localizare temporo-spațială a senzațiilor dureroase. Împreună cu talamusul,
care reprezintă cea mai mare stație senzitivă pentru majoritatea aferențelor,
implicit pentru percepția durerii, ariile corticale (în principal aria
senzitivă I și II) au de asemenea abilitatea de a evalua, integra și interpreta
în final influxul de stimulii dureroși. La ariile somato-senzitive I și II de
proiecție corticală se mai adaugă participarea lobului frontal, a operculumului
lateral și a girusului cingulat anterior, structuri care sunt implicate
împreună cu nucleii talamici dorsomedial și anterior în reacțiile psihologice
la durere. Sunt asociate de asemenea la percepția durerii structuri cerebrale
care primesc prin colaterale ale TSTL aferențe nociceptive: substanța gri
periapeductală, cerebelul, hipotalamusul, amigdala și unii nuclei din
ganglionii bazali.
Tractul
spino-mezencefalic (TSM). TSM este reprezentat de axoni ai neuronilor
nociceptivi de ordinul II din lamina I și V a cornului posterior, care se încrucișează
(comisura albă anterioară), după care se poziționează în partea anterolaterală
a căii, terminându-se în neuronii substanței gri periapeductale, jucând un rol
important în modularea durerii prin activarea unor neurostructuri ce emit
mesaje inhibitorii descendente asupra neuronilor nociceptivi spinali.
Tractul
spino-reticular (TSR) (paleo-spino-talamic). TSR este o parte componentă a
căii spino-reticulo-talamice care se finalizează în neuronii din nucleii
talamici intralaminari, hipotalamus și sistemul limbic. El vehiculează
informații primite de la neuronii laminelor spinale VI, VII și VIII ce răspund
senzitiv de întinse câmpuri receptoare periferice. Această cale este prevăzută
cu multe colaterale și sinapse (cale multisinaptică), fapt care conferă caracterul
difuz al durerii evocate pe această cale, atât somatice, cât și vegetative.
Senzațiile transportate prin TSR sunt integrate în răspunsurile vegetative
reflexe și contribuie la amprenta afectiv-emoțională și motivațională a
comportamentului la durere.
Un câmp receptor cu
importanță aparte în practica clinică este cel al teritoriului cefalic de care
răspunde nervul cranian trigemen. Fața, cavitatea orală, articulația
temporomandibulară, țesuturile cerebrovasculare ale regiunii craniofaciale sunt
frecvent sedii ale durerii în organism, studiile epidemiologice indicând o
prevalență înaltă atât pentru durerea acută, cât și pentru cea cronică (9, 10).
Informațiile nociceptive (durerea de tip „înțepătură”), extracția dentară,
stimulii chimici, lichidele foarte reci sau fierbinți, toxinele bacteriene cu
acces la pulpa expusă sau inflamații orale, temporomandibulare, ale corneei
etc.) sunt receptate de fibre A-delta și C trigeminale reprezentate de dendrite
ale neuronilor din ganglionul trigeminal Gasser (neuroni de ordinul I). Axonii
acestora proiectează în complexul nuclear senzitiv trigeminal format din
nucleul senzitiv principal pontin și nucleul tractului spinal reprezentat de
trei subnuclei: oralis, din treimea inferioară a punții, interpolaris,
din bulb, și caudalis, din măduva cervicală superioară (toți neuroni de
ordinul II). Subnucleul caudalis (numit și cornul posterior al bulbului)
și mai ales neuronii substanței gelatinoase din lamina II a acestuia constituie
releul principal al căii trigemino-talamo-corticale de transmitere a durerilor
din regiunea cefalică. Neuronii de ordinul II realizează conexiuni cu formația
reticulară, nucleul parabrahial și nuclei motori cranieni ce stau la baza
circuitelor din trunchiul cerebral care răspund de răspunsurile reflexe la
stimulii dureroși orofaciali. Neuronii senzitivi trigeminali proiectează după
încrucișarea fibrelor, atât direct, cât și pe căi multisinaptice, în nucleul
ventro-postero-talamic (talamusul ventro-bazal cu neuroni de ordinul III). De
aici fibrele neuronilor talamici ajung în cortexul cingulat anterior și
cortexul somato-senzitiv.
Semnalele dureroase
aferențiale ce parcurg căile enumerate mai sus sunt procesate (analizate,
integrate) și direcționate în final spre cortexul somato-senzitiv. Procesarea
informațiilor (analiza, sistematizarea și codificarea influxurilor
aferențiale) începe chiar din neuronul I (ganglionar), dar prelucrările
neuronale cresc în complexitate pe măsura parcurgerii traseelor ascendente, în
centre importante ca: neuronii cornului posterior și ai nucleilor senzitivi ai
nervilor cranieni (în special nervul trigemen), trunchi cerebral, talamus,
corpii striați și cortexul cerebral. În formațiunile trunchiului cerebral
există relee sinaptice pentru căile algoconductoare ascendente: bulb (nucleii
reticulari, nucleul magnus al rafeului), punte (nucleul parabrahial, locus
coeruleus) și mezencefal (substanța gri periapeductală, formația reticulară
paramediană, nucleul cuneiform, tuberculii quadrigemeni). Talamusul reprezintă
cea mai complexă și importantă stație senzitivă subcorticală de pe traseul
căilor algoconductoare, constituind o arie de proiecție somatotopică și de
organizare care prefigurează în mare măsură proiecția corticală, mergând până
la reprezentarea la nivelul nucleilor talamici a unui homunculus senzitiv
similar cu cel neocortical postcentral.
Procesarea corticală a
durerii are loc în paleo- și neocortex. În sistemul limbic și în strânsă
legătură cu hipotalamusul, este asigurată coloratura afectiv-emoțională individuală
a durerii. Aspecte motivaționale și chiar de control, contribuie la realizarea
fenomenului de „trăire a durerii” de către fiecare subiect în parte, cu răsunet
semnificativ în reacțiile de răspuns comportamental (motorii somatice, motorii
și secretorii vegetative, endocrine, metabolice). Ariile paleocorticale ale
resimțirii și interpretării emoționale a durerii (răspunsul psihologic) sunt:
cortexul prefrontal, cortexul parietal inferior, cortexul cingulat anterior,
insula, amigdala, hipocampul și nucleul accumbens, fiecare cu rolurile sale
destul de bine delimitate în prezent. Neocortexul integrează prin cele trei
arii senzitive – aria somestezică I (girusul postcentral), aria somestezică II
(partea posterioară a marginii superioare a șanțului silvian), aria somestezică
III (suprafața medială a emisferelor cerebrale) – aferențele algogene deja
procesate parțial până aici. Proiecțiile corticale ale durerii în aria I sunt
controlaterale (încrucișate), foarte puțin ipsilaterale și cu reprezentare
inegală a diferitelor zone corporale (homunculus senzitiv). Aria II are o
somatotopie puțin precisă și conexiuni cu talamusul mai restrânse comparativ cu
aria I, fiind implicată esențial în sensibilitatea termică și durerea de tip
„înțepare”. Centrii înalți ai durerii, începând de la talamus, integrează
experiența dureroasă în termeni ce includ intensitatea și localizarea precisă a
durerii, durata, variabilitatea acesteia (de la plăcere până la neplăcere),
aceste atribute făcând din durere mai mult decât o senzație, și anume o
percepție.
Componenta neurochimică
a transmisiei durerii este reprezentată de neurotransmițători din care fac
parte aminoacizi excitatori, un grup numeros de neuropeptide, prostaglandine,
monoxidul de azot, adenozina. Un rol important în transmisia neurochimică
sinaptică îl joacă receptorii membranari neuronali ionotropici de care se leagă
mesagerii primari (neurotransmițătorii durerii). Frecvent, receptorii sunt
asociați unui canal membranar neuronal cationic, cel mai reprezentativ receptor
algogen fiind R-NMDA, care, odată activat prin scoaterea ionului de Mg2+
din interiorul canalului ionic, deschide astfel porul acestuia pentru influxul
de Ca2+ și Na+ asociat cu efluxul de K+.
Sisteme neuromodulatoare ale
durerii
Modularea reprezintă un
proces fiziologic de adaptare și apărare a organismului, o reacție de tip
homeostazic, la capătul căreia are loc instalare unui echilibru în ceea ce
privește percepția durerii în condițiile reducerii sau chiar evitării
tulburărilor neurofuncționale. Baza neuroanatomică a modulării durerii este
dată de circuite scurte și lungi. Din categoria circuitelor scurte fac parte
circuitele inhibitoare post- și presinaptice, circuite recurente și, cel mai
cunoscut, mecanismul controlului de poartă al lui Melzack și Wall. Un rol
important în funcționarea acestor circuite îl au sistemul interneuronilor
inhibitori care modulează activitatea aferențelor dureroase la diferite
niveluri și care a fost bine demonstrată la nivelul releului spinal și al
trunchiului cerebral. Interneuronii inhibitori se găsesc în substanța
gelatinoasă a cornului posterior medular în straturile II și III Rexed, iar
acțiunea lor este tonică și permanent inhibitorie asupra sinapsei fibrelor
A-delta și C cu neuronii nociceptivi din cornul posterior. Circuitele lungi
sunt sisteme descendente sau ascendente care pot inhiba sau facilita tonic și
fazic răspunsul neuronilor nociceptivi. Căile descendente lungi au ca loc de
plecare structuri nervoase supramedulare care sunt stimulate prin colaterale
emise în principal de tractul spino-reticulo-talamic. Cele mai studiate și
cunoscute sunt cele cu origine în substanța gri periapeductală, nucleii
rafeului, formația reticulată, ganglionii bazali (globus pallidus și substanța
nigra), amigdala și hipotalamus. Căile modulatorii au fost denumite în funcție
de natura neuromodulatorului sinaptic, inhibitor/excitator, asupra neuronilor
nociceptivi din lamina I, II, III, IV, V Rexed din substanța gelatinoasă a
cornului posterior cu care fac sinapsă și de asemenea a celor din nucleii
senzitivi trigeminali. Cele mai cunoscute sunt căile descendente inhibitorii
ale durerii, căile opiatergice (folosesc ca neuromodulatori opioidele endogene)
și căile neopioide (serotoninergică, noradrenergică, colinergică, gabaergică).
La nivelul măduvei spinării, majoritatea circuitelor scurte sau lungi au ca
neuromodulatori opioidele și serotonina. Terminațiile sinaptice A-delta și C pe
neuronii nociceptivi ai cornului posterior medular au ca transmițător substanța
și acidul glutamic ca mediatori ai durerii, dar sunt prevăzute și cu receptori
opioizi (miu, delta, kappa) pentru liganzii morfinici proprii (enkefaline,
endorfine, dinorfine) sau exogeni, care depresează eliberarea substanței P
algomediatoare, inhibând invariabil transmisia durerii.
Neuromatrixul durerii și
plasticitatea neuronală
Condițiile existenței
unui tablou fiziologic atât de complex implicat în neurotransmisia și
neuromodularea durerii sunt argumente ce pledează pentru existența unei rețele
neuronale (matrice) la nivelul sistemului nervos periferic și central, cu
efecte atât pronociceptive, cât și antinociceptive, acționând fiecare din ele
prin trei vectori: unul neuroanatomic al căilor și centrilor nervoși, unul
biofizic al câmpurilor bioelectrice excitatoare (depolarizante)/inhibitoare
(hiperpolarizante) și unul neurochimic al neurotransmițătorilor,
neuromodulatorilor și receptorilor cu acțiuni pre- și postsinaptice. Cele trei
interfețe au mecanisme și efecte strâns interrelaționate, interactive,
interdependente și complementare. Răspunsul fiziologic al organismului în fața
durerii este unitar prin cooperarea mecanismelor menționate. Creșterea
intensității, a frecvenței sau a duratei stimulilor dureroși poate induce în
cazul acțiunii lor prelungite la creșterea sensibilității terminațiilor
nervoase periferice, îndeosebi în leziunile mecanice tisulare medii sau în
inflamații. Are loc o sensibilizare periferică prin scăderea pragului de
excitabilitate nociciceptivă, care frecvent poate induce o creștere și a
excitabilității neuronilor centrali (sensibilizare centrală cu scăderea
pragului central de excitabilitate), inclusiv pe linia genelor neuronale
răspunzătoare de sinteza și echilibrul neuropeptidelor
pronociceptive/antinociceptive. Aceste modificări au loc în cadrul proprietății
de neuroplasticitate neuronală ce se activează la neuronii supuși procesului de
convergență a aferențelor nociceptive. Sensibilizarea centrală este la început
dependentă de nivelul de siguranță al barajului sistemelor neuromodulatoare
inhibitoare, dar, odată declanșat, procesul de sensibilizare poate fi preluat,
susținut și reflectat de procesarea disfuncțională a informațiilor nociceptive
(11). Modificările neuroplastice pot să se mențină ore, zile sau chiar
săptămâni depinzând în particular de tipul de agresiune tisulară sau de tipul
de durere acută sau cronică. În clinică, sensibilizarea neuronală centrală, în
care un rol important îl joacă activarea receptorilor de tip R-NMDA și a
receptorului de neurokinină, este un factor esențial pentru dezvoltarea durerii
cronice caracterizată prin hiperalgezie, allodinie, dureri spontane și senzații
dureroase referite în alte organe, altele decât sediul leziunii. Este de
subliniat că sensibilizarea centrală reprezintă totuși un fenomen fiziologic
reversibil (12), ca răspuns la cele mai multe leziuni inflamatorii sau
neinflamatorii.